


In a recent article with Brian Tully, we talked about milk fatty acids and how they relate to feed bunk management – “empty bunk syndrome.” But what was our theory behind using milk fatty acids as a monitor of rumen metabolism? This is an example of serendipity; research can have unexpected spin-offs. How did serendipity play a role in our understanding of how it impacts milk fat synthesis in dairy cows?

The story really begins in 2000, when we began studying the nutritional value of Pima cottonseed in the diet of lactating dairy cows at UC – Davis. The Pima cottonseed we fed to lactating cows was cracked because Pima cottonseed is naturally low in seed coat lint and if the seeds are fed whole they can pass undigested into the feces.
During this feeding study using cracked Pima, a graduate student noticed that there were days when some cows seemed to be low in feed intake. As we feed experimental cows individually, we can notice changes as small as one to two pounds of feed intake per day.
On a commercial dairy, these small differences would likely go undetected, but in a research study these small changes can tell us something about the diet.
We were doing milk FA analysis because of our ongoing focus to modify the FA in milk fat to enhance nutritional value for humans. Scott Taylor, who was performing the FA analysis, noticed that in the milk fat of one cow, there was C18:1 trans 10.
As we rarely, if ever, saw this FA in milk fat from our cows, we spoke with the graduate student and learned that this cow was slightly off-feed.
We initiated our typical treatment for cows that are a little low in feed intake: feeding three to five pounds of long oat hay, and feed intake of this cow was back to normal the next feeding and the C18:1 trans 10 in the cow’s milk fat was gone or at least below our detection limits at that time.
Throughout this study, we had cows slightly off-feed on several days and their milk fat had C18:1 trans 10. That is how we began using C18:1 trans 10 in milk fat as a diagnostic tool to monitor rumen metabolism for our research feeding studies.
So why C18:1 trans 10? This FA is an intermediate in biohydrogenation of unsaturated FA in the rumen. What is biohydrogenation?
Basically it is a process whereby, during normal rumen metabolism, hydrogen is added to the carbon at the site of the double bond, resulting in a single bond. The hydrogenation process also creates trans FA. These trans FA can eventually end up in milk fat.
There are many theories as to why the rumen microbes hydrogenate unsaturated FA, but one thing we know for sure – biohydrogenation is a normal process of rumen metabolism.
Let’s take a step back. There is research going on with milk fat depression and the link to C18:1 trans 10 and another FA, C18:2 trans 10 cis 12. But we are not talking about milk fat depression in the classical sense, where milk fat drops below 3.0 percent and in some research studies, even below 2 percent!
This situation of milk fat depression is often a result of feeding unbalanced rations with too low fiber, too small a fiber length, or too high rumen fermentable carbohydrates.
In California, rations of dairy cattle are generally nutritionally well-balanced and, in the field, a slight decrease in milk fat where the bulk tank milk fat drops from 3.6 percent/3.8 percent to 3.3 percent/3.5 percent is considered to be “milk fat depression.” And this is not related to hot weather – which is a much different scenario than what is occurring with milk fat depression, at least in our opinion.
So these metabolically modest, but financially large, milk fat depressions that we are dealing with occur in cows fed nutritionally well-balanced rations where there is something slightly off in the ration that is perturbing rumen metabolism, so that a shift in the biohydrogenation pathway occurs related to the microbes in the rumen.
When we feed cows, we are actually feeding the microorganisms that live in the rumen, so diet changes will affect microbial populations. Changing the length of the forage in the diet, adding dietary buffers, changing ingredients in the diet – these dietary changes all cause shifts in the microbial population in the rumen.
And the rumen microbial populations are dynamic and change in response to changes in levels of substrate (e.g. starch) in the diet. With more starch in the diet, there will be an increase in the numbers of microbes that utilize starch. These shifts are normal and allow us to feed a wide variety of feedstuffs to dairy cattle that are converted into milk and meat.
However, at times we can cause upsets to rumen metabolism. For example, if we finely grind the forage in the diet, we can cause rumen upsets.
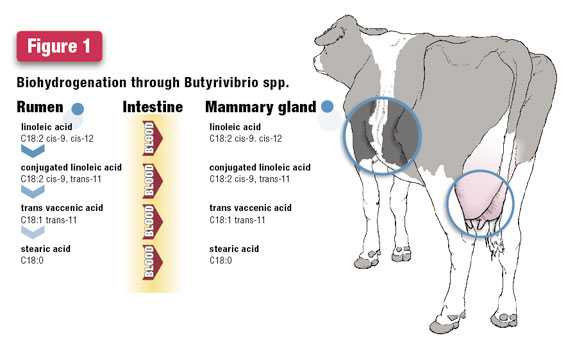
Back to biohydrogenation and rumen metabolism. There is an excellent paper published by researchers at the University of Aberdeen in Scotland, which supports our theory. These researchers showed two proposed pathways that biohydrogenation of C18:1 cis 9 cis 12 can follow in the rumen.
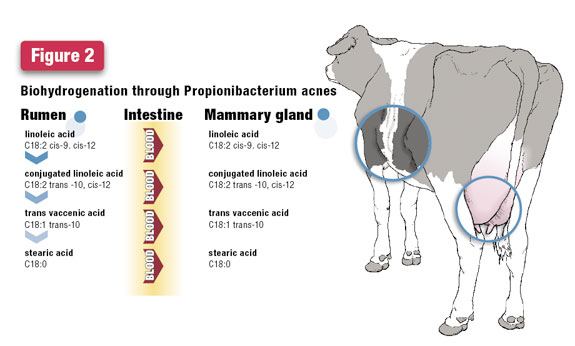
Shifting the pathway from the microbial species Butyrivibrio spp. to Propionibacterium acnes changes the intermediates of rumen biohydrogenation.
The predominant pathway is likely through Butyrivibrio spp. because milk fat typically contains measureable amounts of C18:1 trans 11, but in our research not much C18:1 trans 10 is found.
But in herds with lower milk fat we found C18:1 trans 10, and in one case as high as 2 percent of the total FA. The presence C18:1 trans 10 has been linked to lower milk in published research.
Therefore, our theory is based upon something in the diet shifting the metabolism of biohydrogenation in the rumen from the pathway that yields C18:1 trans 11 to the pathway that yields C18:1 trans 10.
While both pathways are always present, now the pathway that produces C18:1 trans 10 has increased, and this increase in C18:1 trans 10 is linked to lower milk fat percentage.
So what could be slightly off in a ration to affect rumen metabolism? The diets we fed in our cracked Pima rations were nutritionally well-balanced, but there was something slightly off. But what was slightly off?
Our belief is that the lipid in the Pima cottonseed was too rapidly available for biohydrogenation in the rumen. Cottonseed oil is highly unsaturated and contains high amounts of linoleic acid (C18:2 cis 9 cis 12) which, with 2 cis double bonds, is a polyunsaturated FA.
Our thought is that whole cottonseed (i.e. Acala/Upland fuzzy white cottonseed) fed whole to dairy cattle provides cottonseed oil that is slowly released in the rumen so that normal rumen biohydrogenation is not affected.
In contrast, cracking the Pima seed exposes the oil, thereby making the polyunsaturated FA readily available in the rumen so that rumen biohydrogenation is affected. It is not a matter of good or bad. It is simply that metabolism of the unsaturated FA is altered, a natural response of the rumen microorganisms.
However, the impact on production performance may not be what we want. This idea that low milk fat percentage is caused by the relative availability of lipids in the rumen is supported by research conducted in the 1970s in Scotland by Steele, Noble and Moore.
These researchers did some classic research on dietary lipids and milk FA. In one study they compared two methods of adding soybean oil to the diet.
They added the soybean oil as free oil (oil rapidly available to rumen biohdyrogenation) or oil in the form of cracked soybeans (oil slowly available to rumen biohydrogenation).
Cows fed the free soybean oil had higher C18:1 trans 11 in milk fat than cows fed the cracked soybeans.
These shifts in biohydrogenation pathways can be detected by the FA in milk fat. In the normal events of digestion, the FA in the biohydrogenation pathway flow out of the rumen into the small intestine, where they can be absorbed into the blood.
When blood flows to the mammary gland, these FA can be taken up by cells and used to synthesize milk fat. For the pathway through Butyrivibrio spp., we find C18:1 trans 11 in milk fat.
Dietary factors that cause a higher amount of biohydrogenation proceeding through Propionibacterium acnes will be reflected by higher C18:1 trans 10 in milk fat.
What practices might cause a shift in biohydrogenation pathways? Low rumen pH is proposed to shift the pathway from proceeding through the microbial population of Butyrivibrio spp. to the microbial population of Propionibacterium acnes.
How low a rumen pH? We don’t know the answer. The ionophore, Rumensin®, is also thought to shift the pathway through Propionibacterium acnes.
We recently worked with nutritionists in California to analyze bulk tank milk samples from herds that had milk fat between 3.3 and 3.5 percent – slightly lower than normal in the fall, when hot weather was not a likely factor. In these milk fat samples, we found C18:1 trans 10 but no C18:2 trans 10 cis 12.
Thus, based on our theory, we know that something is perturbing rumen metabolism such that the biohydrogenation pathway is shifted to the pathway with Propionibacterium acnes. Now the challenge is to determine what is causing this shift in microbial population.
Using the milk FA data from milk bulk tanks with lower-than-normal milk fat percentage, the nutritionists in the field identified problems and increased milk fat percentage.
Some of the solutions found by a professional nutritionist working on dairy farms included:
- Feedbunk management. This was referred to as “empty bunk syndrome” in a previous article. Empty feed bunks for periods of the day encourage cows to overeat when they are fed. This problem was corrected by ensuring cows always had feed available.
- Reducing distillers grains in the ration. Distillers grains contain a readily available source of dietary lipid that can perturb rumen fermentation. The free oil in distillers is rapidly available in the rumen and can cause a problem in some diets.
- Remove Rumensin®. Ionophores are fed because they affect some microbial populations, which results in increased propionic acid concentration. We know that ionophores can also affect the microbes involved in the biohydrogenation pathways. There is also some speculation that ionophores can exacerbate the effect of high free oil in the rumen.
- Mycotoxins. Moldy feeds contain mycotoxins and these mycotoxins can affect rumen microbes and alter rumen metabolism. One nutritionist reported that removing moldy hominy and corn gluten that were tested and found to contain a mycotoxin increased milk fat percentage back to the herd normal.
- Lysine. There is ongoing research at UC – Davis that unexpectedly found that dietary lysine can impact the fat content of milk – another example of research serendipity! We don’t have all of the information, but there is also information in the literature to support the work at UC – Davis that dietary amino acids can impact milk fat percentage.In this case, the herd with low milk fat did not have C18:1 trans 10 in the bulk tank milk. So lower milk fat percentage does not always have to be directly related to rumen metabolism.
- Others? There will certainly be other factors that will come to light that impact microbes and perturb the metabolic pathways in the rumen that negatively affect milk fat percentage.
So for now, we believe that C18:1 trans 10 in milk fat could be linked to lower milk fat, because it indicates that something in the diet is causing a shift in the biohydrogenation process in the rumen to yield more C18:1 trans 10.
How this higher C18:1 trans 10 affects the synthesis of lipids in the mammary gland is not completely understood, but a number of researchers at various universities are studying the mechanisms involved. You never know when serendipity will strike and benefit the industry. PD

-
Ed DePeters
- Department of Animal Science
- University of California-Davis
- Email Ed DePeters


